Kuwanon T și Sanggenon A izolate din Morus Alba exercită efecte antiinflamatorii prin reglarea căilor de semnalizare NF-κB și HO-1/Nrf2 în celulele BV2 și RAW264.7
Mar 30, 2022
Pentru mai multe informatii. a lua legaturatina.xiang@wecistanche.com
Abstract: Am investigat anterior extractul metanolic din scoarța de Morus alba și am caracterizat 11 compuși din extract: kuwanon G(1), Kuwana E (2), Kuwana T(3), sanggenon A (4), sanggenol M (5), sanggenol A(6), mulberofuran B(7), mulberofuran G(8), moracin M(9), moracin O(10) și norartocarpanonă (11). Aici, am investigatantiinflamatorefectele acestor compuși asupra celulelor microgliale (BV2) și macrofagelor (RAW264.7). Printre acestea, 3 și 4 au inhibat semnificativ producția de oxid nitric indusă de lipopolizaharide (LPS) în aceste celule, sugerând proprietățile antiinflamatorii ale acestor doi compuși. Acești compuși au inhibat producția de prostaglandină E2, interleukină-6 și factor de necroză tumorală-c, precum și expresia oxid nitric sintetazei și ciclooxigenazei-2 inductibile după stimularea LPS. Pretratamentul cu 3 și 4 a inhibat activarea căii de semnalizare a factorului nuclear kappa B în ambele tipuri de celule. Compușii au indus, de asemenea, expresia hemoxigenazei(HO)-1 prin activarea factorului nuclear eritroid 2-factorul 2 înrudit. Suprimarea activității HO-1 a inversat efectele antiinflamatorii cauzate prin pretratament cu 3 și 4, sugerând că efectele antiinflamatorii au fost reglate de HO-1. Luate împreună, 3 și 4 sunt potențiali candidați pentru dezvoltarea agenților terapeutici și preventivi pentruboli inflamatorii.
Cuvinte cheie: Morus alba; kuwanon T; sanggenon A; BV2; celule RAW264.7; efecte antiinflamatorii

Click aici pentru a afla mai multe produse
1. Introducere
Morus alba, o plantă medicinală aparținând familiei Moraceae, a fost folosită pentru tratarea afecțiunilor inflamatorii pulmonare în medicina tradițională [1]. Scoarța sa rădăcină conține diverse componente active, cum ar fi umbeliferon, scopoletin, mucine, taninuri,flavonoide(morusin, mulberrin, mulberrichromene, cyclomulberrin, moracin P, moracin O, mulberrofuran Q, Kuwana E și kuwanon H)[2], 2-arilbenzofurani (moracenin D, moracin P, moracin O și mulberrofuran O), și flavonoide prenilate (licoflavona C, ciclomulberrin, neociclomorusină, sanggenon I, morusin și Kuwana U)[3,4]. Extractele și compușii activi de M. alba pot ameliora bolile pulmonare, după cum au arătat studiile recente [5-7]. Pe lângă efectul farmacologic al M.alba asupra plămânilor, flavonoidele izoprenilate (sanggenol Q, Kuwana T, sanggenon N, mulberrofuran G și mulberrofuran C) exercită efecte hepatoprotectoare în celulele HepG2 induse de t-BHP [8]. Prenil-flavonoidele (kuwanon A, Kuwana C, Kuwana T și morusin) și triterpenoidele (acid betulin, avail și -sitosterol) inhibă substanțial diferențierea adipocitelor 3T3-L1 [9]. Hidratul de Morin, constituentul major al flavonoidului M. alba, atenuează afectarea cronică imprevizibilă a memoriei indusă de stres, indicând faptul că acest compus ar putea stimula sistemul de apărare antioxidantă și ar inhiba căile neuroinflamatorii [10]. Aceste studii sugerează că constituenții conținuti în M.alba sunt potențiali candidați pentru tratarea diferitelor boli.
Inflamaţie, un răspuns complex de autoapărare la stimuli dăunători, joacă un rol important în apărarea imună prin activarea mai multor celule imunitare, inclusiv macrofage, monocite, leucocite, mastocite și alte tipuri de celule [11]. Macrofagele sunt cele mai abundente și mai larg distribuite celule imunitare din organism, iar microglia sunt macrofage rezidente în sistemul nervos central (SNC)[12]. Macrofagele și celulele microgliale sunt jucători cheie în răspunsul inflamator. Ele pot fi activate ca răspuns la stimuli precum lipopolizaharide (LPS), citokine și chemokine și pot induce stări inflamatorii [13]. În condiții inflamatorii, macrofagele și celulele microgliale excesiv de activate determină reglarea anormală a mediatorilor proinflamatori (oxid nitric (NO), prostaglandina E2 (PGE), oxid nitric sintaza inductibilă (iNOS) și ciclooxigenază (COX)-2 ) și citokine proinflamatorii (interleukina (IL)-6 și factorul de necroză tumorală (TNF)-a), prin activarea căii de semnalizare a factorului nuclear kappa B (NF-B)[14,15]. Nivelurile crescute de mediatori proinflamatori exacerbează și mai mult progresia bolilor inflamatorii, care cresc și mai mult activarea factorilor inflamatori, rezultând astfel un cerc vicios[16]. Prin urmare, reglarea mediatorilor inflamatori este esențială pentru tratamentul și prevenirea bolilor inflamatorii.
Hemoxigenaza (HO)-1 este o enzimă limitatoare de viteză care catalizează degradarea hemului în biliverdină, ion feros (Fe2 plus ) și monoxid de carbon (CO)[17]. Inducerea HO-1 este reglată de activarea factorului nuclear eritroid 2-factorul 2(Nrf2). HO-1 poate fi indus ca răspuns la stresul oxidativ și inflamație pentru a proteja țesuturile și a menține homeostazia în organism[18]. Prin urmare, țintirea inducției HO-1 este una dintre strategiile potențiale pentru tratarea bolilor inflamatorii.
În eforturile noastre continue de a explora candidați din produse naturale pentru tratarea bolilor inflamatorii, prezentăm aiciefecte antiinflamatoriide 11l compuși izolați din M.alba în celulele BV2 și RAW264.7 induse de LPS.

2. Rezultate
2.1. Efectele a 11 compuși izolați din M.alba asupra viabilității celulelor BV2 și RAW264.7
În studiul anterior, scoarța de rădăcină de M.alba a fost extrasă în metanol apos, iar extractele obținute au fost împărțite succesiv în EtOAc, n-BuOH și H2O2. Cromatografia repetată pe coloană SiO2, ODS și Sephadex LH-20 a fracției de EtOAc a dat 11 compuși, cum ar fi kuwanon G(1), Kuwana E(2), Kuwana T (3), sanggenon A (4), sanggenon M(5), sanggenol A(6), mulberofuran B(7), mulberofuran G(8), moracin M(9), moracin O(10) și norartocarpanonă (11)[6,19,20]. Structurile chimice a 11 compuși izolați din M.alba sunt ilustrate în Figura 1. Pentru a determina efectele citotoxice ale acestor compuși, un 3-(4,5-dimetiltiazol-2-il){{ 26}}.5-S-a efectuat testul cu bromură de difeniltetrazoliu (MTT), celulele BV2 și RAW264.7 au fost tratate cu concentrațiile indicate de compuși timp de 48 de ore, la concentrații de până la 80 μM. (Figura 2). Compușii 2, 3 și 4 au avut efecte toxice la 80 μM, compusul 5 a avut efecte toxice la 40 μM. Pe baza rezultatului evaluării toxicității, a fost selectat un interval de concentrație netoxic pentru studiile ulterioare privind efectele antiinflamatorii (compusul 2, 3,4 la 40 uM, compusul 5 la 200 μM și alți compuși la 80 μM) .

 and RAW264.7 (B) cells. The cells were incubated for 48 h with various concentrations of the compounds, and their viability was determined using MTT (3-(4,5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay. Error bars represent mean ± standard deviation of three independent experiments. * p < 0.05, ** p < 0.01 ,*** p < 0.001 compared with the control group. Figure 2. Cytotoxic effects of compounds 1–11 isolated from M. alba on BV2 (A) and RAW")
2.2.Efectele compușilor 11 izolați din M. alba asupra expresiei factorilor inflamatori și proteinei iNOS în celulele BV2 și RAW264.7
NO este un radical liber în sistemul cardiovascular, nervos șisisteme imunitare. Menține homeostazia intracelulară, transportă neurotransmițători și reglează activitatea antiinflamatoare și citotoxicitatea. Cu toate acestea, atunci când se produce o cantitate mare de NO, acesta are efecte dăunătoare asupra organismului, inclusiv vasodilatație, citotoxicitate și leziuni tisulare [21,22]. Apoi am examinat efectele acestor compuși asupra producției de nitriți în celulele BV2 și RAW264.7 induse de LPS. Celulele au fost tratate cu diferite concentrații de compuși timp de 2 ore înainte de stimularea cu LPS (1 ug/mL) timp de 24 de ore. Dintre compuși, numai Compusul 3 (kuwanonT) și 4 (sanggenon A) au inhibat semnificativ producția de nitriți atât în liniile celulare BV2, cât și RAW264.7 (Figura 3). În plus, am efectuat un experiment suplimentar pentru a compara efectul inhibitor al producției de nitriți între grupul tratat cu LPS după pretratarea compusului și grupul tratat cu compus după pretratarea cu LPS. Ca urmare, nu a existat nicio diferență în ceea ce privește efectul inhibitor al producției de nitriți între cele două grupuri experimentale (Figura suplimentară S1). Prin urmare, în acest studiu, următoarele experimente au fost efectuate folosind pretratarea cu compuși în decurs de 2 ~ 3 ore înainte de tratarea LPS.
 and RAW264.7 (B) cells. The cells were pretreated for 2 h with concentrations of compounds and stimulated for 24 h with lipopolysaccharide (LPS; 1 µg/mL). Error bars represent mean ± standard deviation of three independent experiments. * p < 0.05 and *** p < 0.001 compared with the LPS-treated group")
Producția de NO este crescută de proteinele proinflamatorii inductibile de oxid nitric sintaza (iNOS). Cu toate acestea, supraexprimarea iNOS afectează serios fiziopatologia bolii [23]. Compușii 3 și 4 au inhibat expresia iNOS într-o manieră dependentă de concentrație (Figura 4). Am examinat efectele compușilor 3 și 4 asupra expresiei induse de LPS a factorilor inflamatori în celulele BV2 și RAW264.7. Celulele au fost tratate cu diferite concentrații de compuși 3 și 4 timp de 2 ore înainte de stimularea cu LPS (1 ug/mL) timp de 24 de ore. Ambii compuși au inhibat semnificativ expresia indusă de LPS a PGE2, TNF și IL-6 în celulele BV2 și RAW264.7 (Figura 5). Rezultatele au părut să arate că pre-tratamentul cu compusul 3 și 4 a suprimat inflamația indusă de LPS în celulele BV2 și RAW264.7.


")
2.3. Efectele compușilor 3 și 4 asupra translocării NF-xB în celulele BV2 și RAW264.7
NF-kB este un factor de transcripție care reglează expresia iNOS [24]. NF-kB normal rămâne în forma sa inactivă prin formarea de complecși cu proteine de reglare, cum ar fi IKB. Cu toate acestea, atunci când este activat de LPS, IkBa este degradat prin fosforilare, iar NF-kB (cum ar fi p65) este translocat în nucleu [25], care promovează gena mediatoare inflamatorie și induce expresia factorilor inflamatori [26]. Pentru a examina în continuare efectul inhibitor al compușilor 3 și 4 asupra producției de factori proinflamatori în celulele BV2 și RAW264.7 activate de LPS, am investigat translocarea nucleară a expresiei p65 în celulele tratate cu compușii 3 și 4, LPS sau ambele folosind un anticorp marcat cu FITC anti-p65. DAPI a fost folosit pentru colorarea nucleară. În grupul de control, expresia p65 a fost detectată în citosol. Cu toate acestea, în celulele induse de LPS, acumularea p65 a fost detectată în nucleu, așa cum este indicat în imaginile îmbinate ale colorării DAPI și p65. În plus, în comparație cu celulele induse de LPS, compușii 3 și 4 au redus semnificativ creșterea mediată de LPS a activității de legare a ADN-ului NF-KB (p65) (Figura 6A, B) și translocarea nucleară (Figura 6C-F). Aceste descoperiri indică faptul că compușii 3 și 4 sunt regulatori negativi ai translocației nucleare NF-kB stimulate de LPS.
 and NF-κB (p65) localization (C–F) in BV2 and RAW264.7 cells. The cells were pretreated with compounds 3 or 4 for 2 h and stimulated with liposaccharide (LPS; 1 µg/mL) for 1 h. Experiments were performed using a commercially available enzyme-linked immunosorbent assay kit, as described in the Materials and Methods section. ** p and *** p < 0.001 compared with the LPS-treated group")
2.4. Efectele compușilor 3 și 4 asupra căii Nrf2/HO-1 în BV2 și RAW264.7
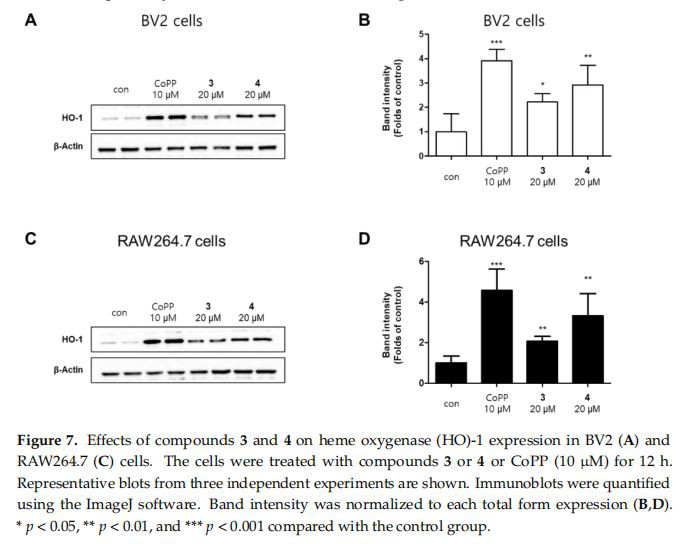
Celulele Hemoxigenaza-1(HO-1) este o țintă a factorului nuclear E2-factorul 2(Nrf2), calea Nrf2/HO-1 este un puternic sistem de semnalizare antioxidant pentru promovarea fierului liber în monoxid de carbon (CO), biliverdină și hem [27]. Monoxidul de carbon, un metabolit gazos al catabolismului hemului, prezintă efecte reglatoare ale vasodilatației și răspunsurilor proinflamatorii [28]. În plus, HO-1 este, de asemenea, cunoscut că joacă un rol important în protejarea celulelor împotriva inflamației și a stresului oxidativ și în reglarea producției de nitriți în macrofagele activate [29]. Am efectuat Western blotting pentru a investiga dacă compușii 3 și 4 măresc nivelul de expresie al HO-1, în care binecunoscutul inductor de HO-1 cobalt protoporfirina (CoPP) a fost utilizat ca control pozitiv pentru creșterea HO{{ 16}} expresia proteinei. [30]. Rezultatele au arătat că compușii 3 și 4 au reglat, de asemenea, expresia HO-1 (Figura 7). Am explorat în continuare dacă compușii 3 și 4 reglează activarea Nrf2. Translocarea Nrf2 la nucleu a fost crescută într-o manieră dependentă de timp, indicând faptul că compușii 3 și 4 au reglat semnificativ calea Nrf2/HO-1 în celulele BV2 și RAW264.7 (Figura 8).
 and RAW264.7 (B,D) cells. The cells were treated with compounds 3 or 4 for 0.5, 1, and 1.5 h. Representative blots from three independent experiments are shown. Immunoblots were quantified using the ImageJ software. Band intensity was normalized to each β-actin or PCNA. * p < 0.05, ** p < 0.01, *** p < 0.001, # p < 0.05, ## p < 0.01, and ### p < 0.001, compared with the control group")
 and RAW264.7 (B,D) cells. The cells were treated with compounds 3 or 4 for 0.5, 1, and 1.5 h. Representative blots from three independent experiments are shown. Immunoblots were quantified using the ImageJ software. Band intensity was normalized to each β-actin or PCNA. * p < 0.05, ** p < 0.01, *** p < 0.001, # p < 0.05, ## p < 0.01, and ### p < 0.001, compared with the control group")
Pentru a examina în continuare dacă efectele anti-neuroinflamatorii și anti-inflamatorii ale compușilor 3 și 4 sunt corelate cu expresia HO-1 în celulele BV2 și RAW264.7, am efectuat un set de experimente cu staniu protoporfirina-IX (SNPP) care este un inhibitor selectiv al activității HO-1. Folosind SNPP care poate regla în jos expresia HO-1, am încercat să determinăm dacă HO-1 mediază efectul inhibitor al compușilor de mai sus asupra răspunsului inflamator indus de LPS 【31】. După tratarea celulelor cu 20 μM de compuși 3 și 4 timp de 2 ore cu sau fără SNPP 50 uM, acestea au fost tratate cu LPS timp de 24 de ore. Deși compușii 3 și 4 au redus producția de nitriți în celulele BV2 și RAW264.7 induse de LPS, efectele lor antiinflamatorii au fost inversate prin tratamentul SNPP (Figura 9). SNPP singur nu a afectat producția de oxid nitric (NO) după stimularea LPS, sugerând că efectele antiinflamatorii ale compușilor 3 și 4 sunt reglate de expresia HO-1.

 and RAW264.7 (B,D,F) cells. The cells were treated with 50 µM of tin protoporphyrin-IX (SnPP) or compounds 3 or 4 and stimulated for 24 h with lipopolysaccharide (LPS; 1 µg/mL). Data are presented as mean ± standard deviation of three independent experiments. ** p < 0.01 and *** p < 0.001")
3. Discuție
Prezentul studiu a demonstrat că Kuwana T și sanggenon A exercită inflamația indusă de LPS suprimată în celulele BV2 și RAW264.7. Acești compuși au inhibat producția indusă de LPS de NO, PGE2 și citokine proinflamatorii, inclusiv IL-6 și TNF- și expresia iNOS și COX-2 în celulele BV2 și RAW264.7. Aceste rezultate au indicat că Kuwana T și sanggenon A și-au exercitat efectele antiinflamatorii prin inactivarea căii de semnalizare NF-kB. În plus, acești compuși au indus expresia HO-1 prin activarea Nrf2. În plus, acești compuși au indus expresia HO{-1 prin activarea Nrf2 și s-a confirmat, de asemenea, că acest efect este legat de activitate antiinflamatoare.
Nitric oxide synthase (NOS) has three isoforms: neuronal NOS (nNOS; NOS1), iNOS (NOS2), and endothelial eNOS (NOS3)[32]. iNOS is an inducible form that is upregulated in response to various stimuli, including LPS, cytokines, chemokines, and stress, while nNOS and eNOS are constitutive forms that catalyze continuous NO secretion at basal concentrations J33]. Similar to iNOS, COX-2 is also an inducible form upregulated by various inflammatory stimuli, such as cytokines, growth factors, tumor promoters, and bacterial LPS[34]. Its other isoform, COX-1, is constitutively expressed in most tissues under normal physiological conditions [35].COX-2 exerts an enzymatic effect on the conversion of prostaglandin H2 (PGH2), which converts arachidonic acid to PGE, prostaglandin 12(PGI2), prostaglandin F2a (PGF>.), și tromboxan A2(TXA,)[361. În condiții inflamatorii, nivelurile de iNOS și COX-2 cresc, ducând la supraproducția de NO și respectiv PGE2 și conducând la exacerbarea tulburărilor inflamatorii [37]. În studiul de față, am evaluat mai întâi 11 compuși izolați din M.alba pentru a determina dacă producția de NO este inhibată în celulele BV2 și RAW264.7 stimulate cu LPS. Rezultatele noastre au arătat că kuwanon T și sanggenon A au exercitat cele mai puternice efecte inhibitoare atât în celulele BV2, cât și în celulele RAW264.7 (Figura 3). În studiul nostru anterior, moracina M a avut un efect antiinflamator asupra macrofagelor alveolare, dar rezultatele noastre au arătat un alt efect. model. Cu toate acestea, deoarece fiecare celulă are caracteristici diferite, pot apărea activități diferite chiar și cu același mecanism J38]. Prin urmare, ca și în rezultatele acestei lucrări, s-a confirmat că moracina M nu a avut activitate antiinflamatoare în celulele BV2 și RAW264.7 până la concentrația de tratament. Prin urmare, kuwanon T și sanggenon A au fost selectați pentru experimente ulterioare.
Citokinele au o influență reglatoare complexă asupra răspunsurilor inflamatorii și imune [39]. Activarea macrofagelor și a celulelor microgliale mărește secreția de citokine proinflamatorii J40, iar această secreție crescută duce la eliberarea ulterioară a citokinelor [41]. IL-6 este una dintre citokinele majore și este un mediator solubil responsabil de inflamație, răspuns imun și hematopoieză [42]. Semnalizarea IL-6 este reglată prin două mecanisme diferite, inclusiv legarea la receptorul IL-6 legat de membrană (mblL6R) și recunoașterea receptorului IL-6 solubil (sIL6R)[43]. Ambele mecanisme sunt asociate cu activarea glicoproteinei (GP)130, care are ca rezultat activarea moleculelor de semnalizare din aval, inclusiv kinaza Janus (IAK)/transductorul de semnal și activatorul kinazelor de transcripție (STAT), kinaza fosfoinozitid 3-(PI3K). ), și protein kinaza activată de mitogen (MAPK) [44]. IL-6 este utilizat pentru a prezice și a evalua nivelurile de inflamație la pacienții cu cancer, infecții, boli autoimune, boli pancreatice și boli cardiovasculare.
TNF-, o altă citokină majoră, joacă, de asemenea, un rol important în multe procese imunitare și inflamatorii, cum ar fi proliferarea celulelor imune, apoptoza, necroza și supraviețuirea. Efectele biologice ale TNF- sunt reglate prin legarea la doi receptori diferiți, receptorul factorului de necroză tumorală (TNFR)1 și TNFR2. TNFR1 este principalul mediator al activității TNF- în majoritatea celulelor, deoarece TNFR2 are o afinitate mai mică de legare pentru TNF-, ceea ce duce la disocierea TNF- mai ușor de TNFR2 decât de TNFR1. Anomalii în semnalizarea TNF și supraproducția de TNF- duc la dezvoltarea multor boli, inclusiv artrita reumatoidă, psoriazisul, boala Crohn, ateroscleroza, sepsisul, diabetul și obezitatea [45,46. Prin urmare, este important să se inhibe producția de citokine proinflamatorii pentru a suprima și a preveni reacțiile inflamatorii și dezvoltarea bolilor inflamatorii. În studiul de față, kuwanon T și sanggenon A au inhibat producția indusă de LPS de IL-6 și TNF- atât în celulele BV2, cât și în celulele RAW264.7.
NF-KB este cel mai omniprezent factor de transcripție[47], iar semnalizarea NF-kB joacă un rol important în exprimarea genelor legate de răspunsurile inflamatorii, inclusiv cele care codifică iNOS, COX-2 și citokinele pro-inflamatorii. 48]. În starea inactivată, subunitățile NF-kB există în citoplasmă legate de proteina sa inhibitoare, inhibitorul kappa B (kB)-. Mai mulți stimuli, inclusiv LPS și citokine, induc fosforilarea și degradarea IkB-, permițând eliberarea și translocarea subunităților NF-kB în nucleu. Subunitățile translocate se leagă de situsurile kB ale genelor țintă din ADN, rezultând inducerea transcripției genelor care codifică mediatori proinflamatori [49]. Prin urmare, inactivarea căii NF-kB poate fi o țintă terapeutică pentru bolile inflamatorii. În studiul de față, pretratamentul cu kuwanon T și sanggenon A a inhibat activarea indusă de LPS a semnalizării NF-kB prin inhibarea activității de legare a ADN-ului și a translocației nucleare a subunităților NF-kB (Figura 6). Aceste rezultate sugerează că efectele antiinflamatorii ale kuwanon T și sanggenon A sunt exercitate prin reglarea căii de semnalizare NF-kB.
În condiții normale, Nrf2 se leagă de proteina 1 (Keap1) asociată ECH-like Kelch din citoplasmă. Cu toate acestea, se disociază de Keapl, se translocă în nucleu și se leagă de situsurile elementului de răspuns antioxidant (ARE) de pe ADN, ducând la exprimarea diferitelor gene antioxidante, inclusiv a celor care codifică HO-1, NAD(P)H : chinona oxidoreductaza 1(NQO1), peroxiredoxină (Prx) și tioredoxină (Trx)[50]. În special, se știe că HO-1 este asociat cu efecte antiinflamatorii, iar activitatea sa este reprezentată de nivelurile de CO, unul dintre cei trei subproduși generați din hem de activitatea enzimatică a HO{{12 }} [51]. În studiul de față, am constatat că kuwanon T și sanggenon A au indus expresia HO-1 prin activarea Nrf2 (Figurile 7 și 8). În plus, am verificat corelația dintre efectul antiinflamator al kuwanonului T și al expresiei sanggenon A și HO-1 folosind SNPP, un inhibitor selectiv al activității HO-1. Efectele inhibitoare ale kuwanon T și sanggenon A asupra producției de NO și TNF și activării NF-kB au fost parțial inversate prin co-tratament cu SNPP (Figura 9). Aceste rezultate sugerează că efectele antiinflamatorii ale kuwanonului T și ale sanggenonului A sunt reglate de expresia HO-1.
")
4. Materiale și Metode
4.1. Materiale
Roswell Park Memorial Institute 1640 (RPMI 1640) și serul fetal bovin au fost achiziționate de la Gibco BRL Co. (Grand Island, NY, SUA). Toate substanțele chimice au fost obținute de la Sigma-Aldrich Chemical Co. (St. Louis, MO, SUA). Anticorpii primari anti-iNOS, anti- -actină, anti-p65 și anti-HO-1 au fost achiziționați de la Santa Cruz Biotechnology (Santa Cruz, CA, SUA); și anticorpi secundari anti-iepure și anti-șoarece de la Millipore
(Billerica, MA, SUA). Trusele de testare imunosorbantă legată de enzime (ELISA) pentru PGE,, IL-6 și TNF- au fost achiziționate de la R&DSystems Inc. (Minneapolis, MN, SUA). Izolarea și determinarea structurală a celor 11 compuși din Morus alba a fost descrisă în altă parte [19-21].
4.2. Cultură celulară și teste de viabilitate
Celulele BV2 și RAW264.7 au fost însămânțate la o densitate de 5 x 105 celule/ml în RPMI 1640 suplimentat cu 1% antibiotic (penicilină-streptomicina) și 10% FBS inactivat la căldură. Celulele au fost cultivate la 37 de grade într-un CO umidificat de 5%, cu o atmosferă de aer de 95%, conform unei metode descrise anterior [52].
4.3. Măsurarea producției de NO
Producția de nitriți, un produs final stabil al oxidării NO, a fost măsurată ca un indicator al producției de NO în celule. Pe scurt, concentrația de nitriți în mediul condiționat a fost determinată folosind o metodă bazată pe reacția Griess [53]. Detaliile testului au fost descrise anterior [54].
4.4. PGE2 Ass4y/
Concentrația de PGEz din fiecare probă a fost măsurată folosind un kit ELISA disponibil comercial, conform unei metode descrise anterior [55].
4.5.Măsurarea nivelurilor IL-6 și TNF-a
Mediul de cultură a fost colectat pentru a determina nivelurile de IL-6 și TNF- folosind un kit disponibil comercial (BioLegend, San Diego, CA, SUA). Testul a fost efectuat conform instrucțiunilor producătorului. Pe scurt, celulele BV2 și RAW264.7 au fost însămânțate în plăci de cultură 24-godeuri la o densitate de 20 × 10 celule/godeu. După incubare, supernatantul a fost colectat și utilizat pentru a măsura concentrațiile de IL-6 și TNF- cu truse ELISA.
4.6. Analiza Western Blot
Celulele peletate BV2 și RAW264.7 au fost spălate cu PBS și lizate în tampon RIPA. Au fost cuantificate cantități egale de proteine utilizând concentrat de reactiv colorant pentru testarea proteinei obținut de la Bio-Rad Laboratories (#5000006; Hercules, CA, SUA), amestecat în tamponul de încărcare a probei și separat folosind SDS-PAGE. Proteinele separate au fost apoi transferate pe o membrană de nitroceluloză. Legarea nespecifică de membrană a fost blocată prin incubare într-o soluție de lapte degresat. Membrana a fost incubată cu anticorpi primari (toți au fost utilizați la 1:1000) la 4 grade peste noapte și apoi a reacționat cu un anticorp secundar conjugat cu peroxidază de hrean (toți au fost utilizați la 1:5000) (Millipore) [ 31].
4.7.Localizarea NF-xB și analiza imunofluorescenței
Pentru a studia localizarea celulelor NF-kB, BV2 și RAW264.7 au fost cultivate pe lame de cameră Lab-Tek II și tratate cu diferite concentrații de compuși timp de 2 ore înainte de stimularea LPS (0.5 ug/mL) timp de 1 h. Celulele au fost apoi fixate în formol și permeabilizate cu acetonă rece și testate cu anticorpi anti-p65 (1:200), urmată de incubare cu un anticorp secundar marcat cu izotiocianat de fluoresceină (1:1000) (Alexa Fluor 488, Invitrogen). Pentru a vizualiza nucleele, celulele au fost tratate cu 1 ug/mL de 4'{6-diamidino{-2-fenilindol (DAPI timp de 30 de minute, spălate cu PBS timp de 5 minute și tratate cu 50 μL VectaShield (Vector Laboratories, Burlingame, CA, SUA). Celulele colorate au fost vizualizate și imaginile au fost achiziționate folosind un microscop cu fluorescență Zeiss (Provis AX70; Olympus Optical Co., Tokyo, Japonia). [31].
4.8. Analize statistice
Datele sunt prezentate ca medie ± abatere standard a trei experimente independente. Analiza unidirecțională a varianței, urmată de testul de comparație multiplă al lui Tukey, a fost utilizată pentru a compara diferențele dintre cele trei grupuri. Analizele statistice au fost efectuate folosind software-ul GraphPad Prism (versiunea 5.01, GraphPad Software Inc., San Diego, CA, SUA),
5. Concluzii
Kuwanon T și sanggenon A au exercitat efecte antiinflamatorii în celulele BV2 și RAW264.7 induse de LPS prin inhibarea producției de NO, PGE2, IL-6 și TNF- și expresia iNOS și COX{{8} }. Rezultatele noastre au demonstrat că aceste efecte inhibitoare au fost mediate prin inactivarea căii NF-KB prin tratamentul cu kuwanon T și sanggenon A. În plus, acești compuși au indus expresia HO-1 prin activarea translocației Nrf2 în nucleu. . Descoperirile noastre au demonstrat că expresia HO-1 indusă de kuwanon T și sanggenon A a fost asociată cu efectele supresive împotriva inflamației induse de LPS. Luate împreună, rezultatele noastre oferă dovezi că Kuwana T și sanggenon A izolate din M. Albu ar putea fi candidați pentru dezvoltarea agenților terapeutici și preventivi pentru boli neuroinflamatorii, cum ar fi boala Alzheimer și boala Parkinson.





